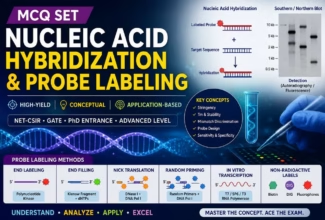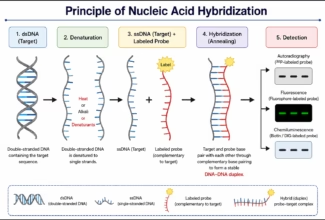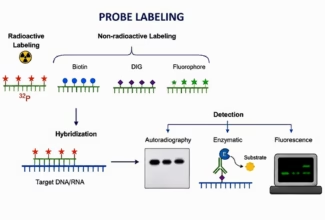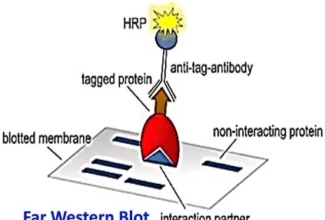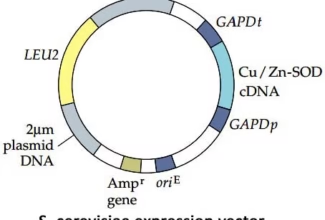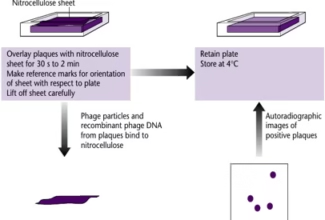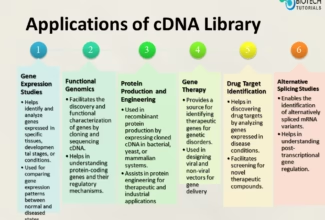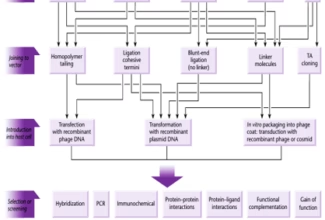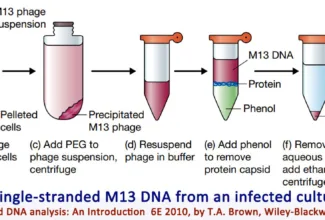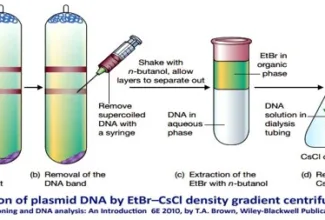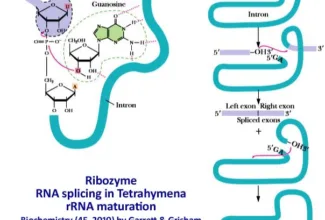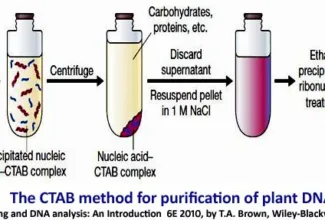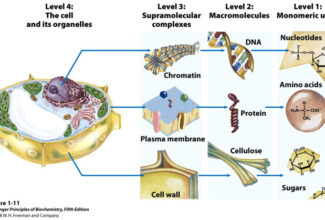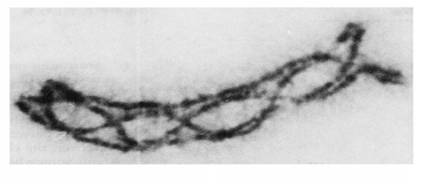
Homologous Recombination (Holliday model and DNA double strand repair model) and Site Specific Recombination
Recombination
- Cleavage and rejoining of DNA molecules to generate new combinations of genes
- There are two types of recombination mechanisms that occur in living system
Genetic recombination
Site specific recombination
Genetic Recombination
- The production of gene combinations not found in the parents by the crossing over between homologous chromosomes during meiosis is called genetic recombination.
- Genetic recombination occurs in the germ cells (during meiosis I in oocytes and spermatocytes)
- Also called as homologous recombination as it takes place between homologous chromosomes
- It occurs during crossing over, in which homologous regions of chromosomes are exchanged and genes are shuffled into new combinations.
- It is an extremely important genetic process because it increases genetic variation.
- It is a ubiquitous process that occurs in every bacteria and also in eukaryotes
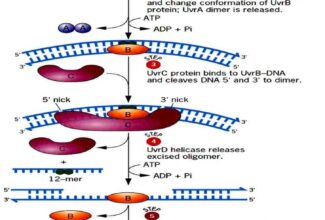
Models of Homologous Recombination
Holliday model (Robin Holliday 1964)
Double strand repair model